
Animal Diversity
14.5 – Birds
Learning Objectives
By the end of this section, you will be able to do the following:
- Describe the evolutionary history of birds
- Describe the derived characteristics in birds that facilitate flight
With over 10,000 identified species, the birds are the most speciose of the land vertebrate classes. Abundant research has shown that birds are really an extant clade that evolved from maniraptoran theropod dinosaurs about 150 million years ago. Thus, even though the most obvious characteristic that seems to set birds apart from other extant vertebrates is the presence of feathers, we now know that feathers probably appeared in the common ancestor of both ornithischian and saurischian lineages of dinosaurs. Feathers in these clades are also homologous to reptilian scales and mammalian hair, according to the most recent research. While the wings of vertebrates like bats function without feathers, birds rely on feathers, and wings, along with other modifications of body structure and physiology, for flight, as we shall see.
Characteristics of Birds
Birds are endothermic, and more specifically, homeothermic—meaning that they usually maintain an elevated and constant body temperature, which is significantly above the average body temperature of most mammals. This is, in part, due to the fact that active flight—especially the hovering skills of birds such as hummingbirds—requires enormous amounts of energy, which in turn necessitates a high metabolic rate. Like mammals (which are also endothermic and homeothermic and covered with an insulating pelage), birds have several different types of feathers that together keep “heat” (infrared energy) within the core of the body, away from the surface where it can be lost by radiation and convection to the environment.
Modern birds produce two main types of feathers: contour feathers and down feathers. Contour feathers have a number of parallel barbs that branch from a central shaft. The barbs in turn have microscopic branches called barbules that are linked together by minute hooks, making the vane of a feather a strong, flexible, and uninterrupted surface. In contrast, the barbules of down feathers do not interlock, making these feathers especially good for insulation, trapping air in spaces between the loose, interlocking barbules of adjacent feathers to decrease the rate of heat loss by convection and radiation. Certain parts of a bird’s body are covered in down feathers, and the base of other feathers has a downy portion, whereas newly hatched birds are covered almost entirely in down, which serves as an excellent coat of insulation, increasing the thermal boundary layer between the skin and the outside environment.
Feathers not only provide insulation, but also allow for flight, producing the lift and thrust necessary for flying birds to become and stay airborne. The feathers on a wing are flexible, so the feathers at the end of the wing separate as air moves over them, reducing the drag on the wing. Flight feathers are also asymmetrical and curved, so that air flowing over them generates lift. Two types of flight feathers are found on the wings, primary feathers and secondary feathers ((Figure)). Primary feathers are located at the tip of the wing and provide thrust as the bird moves its wings downward, using the pectoralis major muscles. Secondary feathers are located closer to the body, in the forearm portion of the wing, and provide lift. In contrast to primary and secondary feathers, contour feathers are found on the body, where they help reduce form drag produced by wind resistance against the body during flight. They create a smooth, aerodynamic surface so that air moves swiftly over the bird’s body, preventing turbulence and creating ideal aerodynamic conditions for efficient flight.
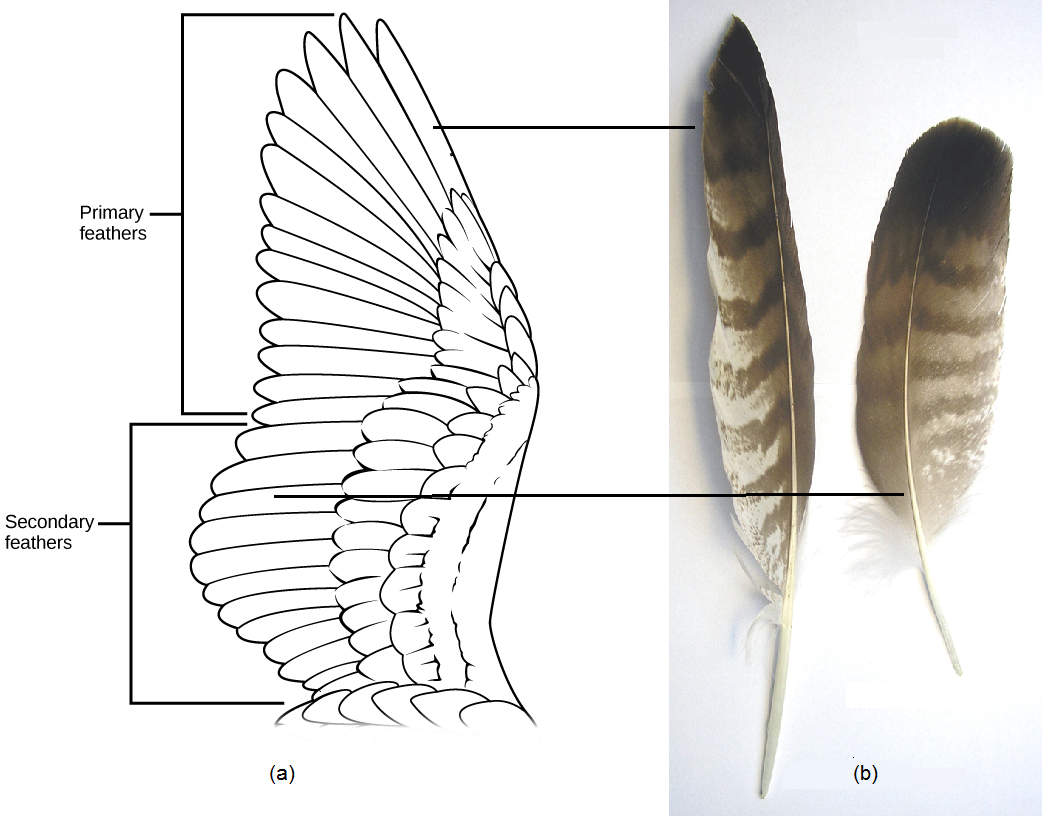
Flapping of the entire wing occurs primarily through the actions of the chest muscles: Specifically, the contraction of the pectoralis major muscles moves the wings downward (downstroke), whereas contraction of the supracoracoideus muscles moves the wings upward (upstroke) via a tough tendon that passes over the coracoid bone and the top of the humerus. Both muscles are attached to the keel of the sternum, and these are the muscles that humans eat on holidays (this is why the back of the bird offers little meat!). These muscles are highly developed in birds and account for a higher percentage of body mass than in most mammals. The flight muscles attach to a blade-shaped keel projecting ventrally from the sternum, like the keel of a boat. The sternum of birds is deeper than that of other vertebrates, which accommodates the large flight muscles. The flight muscles of birds who are active flyers are rich with oxygen-storing myoglobin. Another skeletal modification found in most birds is the fusion of the two clavicles (collarbones), forming the furcula or wishbone. The furcula is flexible enough to bend and provide support to the shoulder girdle during flapping.
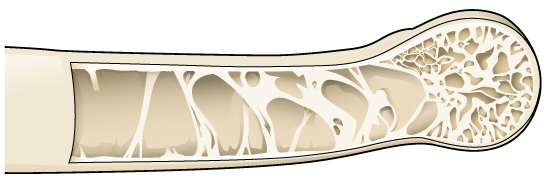
An important requirement for flight is a low body weight. As body weight increases, the muscle output required for flying increases. The largest living bird is the ostrich, and while it is much smaller than the largest mammals, it is secondarily flightless. For birds that do fly, reduction in body weight makes flight easier. Several modifications are found in birds to reduce body weight, including pneumatization of bones. Pneumatic bones ((Figure)) are bones that are hollow, rather than filled with tissue; cross struts of bone called trabeculae provide structural reinforcement. Pneumatic bones are not found in all birds, and they are more extensive in large birds than in small birds. Not all bones of the skeleton are pneumatic, although the skulls of almost all birds are. The jaw is also lightened by the replacement of heavy jawbones and teeth with a beak made of keratin (just as hair, scales, and feathers are).
Other modifications that reduce weight include the lack of a urinary bladder. Birds possess a cloaca, an external body cavity into which the intestinal, urinary, and genital orifices empty in reptiles, birds, and the monotreme mammals. The cloaca allows water to be reabsorbed from waste back into the bloodstream. Thus, uric acid is not eliminated as a liquid but is concentrated into urate salts, which are expelled along with fecal matter. In this way, water is not held in a urinary bladder, which would increase body weight. In addition, the females of most bird species only possess one functional (left) ovary rather than two, further reducing body mass.
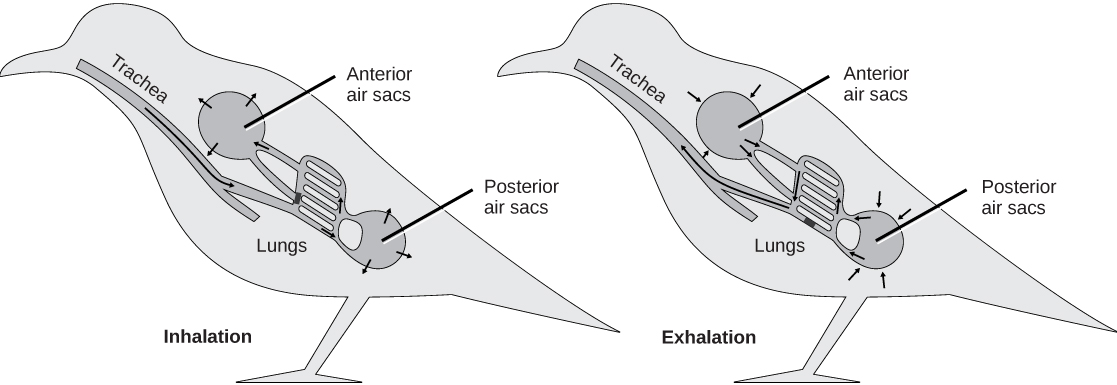
The respiratory system of birds is dramatically different from that of reptiles and mammals, and is well adapted for the high metabolic rate required for flight. To begin, the air spaces of pneumatic bone are sometimes connected to air sacs in the body cavity, which replace coelomic fluid and also lighten the body. These air sacs are also connected to the path of airflow through the bird’s body, and function in respiration. Unlike mammalian lungs in which air flows in two directions, as it is breathed in and out, diluting the concentration of oxygen, airflow through bird lungs is unidirectional ((Figure)). Gas exchange occurs in “air capillaries” or microscopic air passages within the lungs. The arrangement of air capillaries in the lungs creates a counter-current exchange system with the pulmonary blood. In a counter-current system, the air flows in one direction and the blood flows in the opposite direction, producing a favorable diffusion gradient and creating an efficient means of gas exchange. This very effective oxygen-delivery system of birds supports their higher metabolic activity. In effect, ventilation is provided by the parabronchi (minimally expandible lungs) with thin air sacs located among the visceral organs and the skeleton. A syrinx (voice box) resides near the junction of the trachea and bronchi. The syrinx, however, is not homologous to the mammalian larynx, which resides within the upper part of the trachea.
Beyond the unique characteristics discussed above, birds are also unusual vertebrates because of a number of other features. First, they typically have an elongate (very “dinosaurian”) S-shaped neck, but a short tail or pygostyle, produced from the fusion of the caudal vertebrae. Unlike mammals, birds have only one occipital condyle, allowing them extensive movement of the head and neck. They also have a very thin epidermis without sweat glands, and a specialized uropygial gland or sebaceous “preening gland” found at the dorsal base of the tail. This gland is an essential to preening (a virtually continuous activity) in most birds because it produces an oily substance that birds use to help waterproof their feathers as well as keep them flexible for flight. A number of birds, such as pigeons, parrots, hawks, and owls, lack a uropygial gland but have specialized feathers that “disintegrate” into a powdery down, which serves the same purpose as the oils of the uropygial gland.
Like mammals, birds have 12 pairs of cranial nerves, and a very large cerebellum and optic lobes, but only a single bone in the middle ear called the columella (the stapes in mammals). They have a closed circulatory system with two atria and two ventricles, but rather than a “left-bending” aortic arch like that of mammals, they have a “right-bending” aortic arch, and nucleated red blood cells (unlike the enucleated red blood cells of mammals).
All these unique and highly derived characteristics make birds one of the most conspicuous and successful groups of vertebrate animals, filling a range of ecological niches, and ranging in size from the tiny bee hummingbird of Cuba (about 2 grams) to the ostrich (about 140,000 grams). Their large brains, keen senses, and the abilities of many species to imitate vocalization and use tools make them some of the most intelligent vertebrates on Earth.
Evolution of Birds
Thanks to amazing new fossil discoveries in China, the evolutionary history of birds has become clearer, even though bird bones do not fossilize as well as those of other vertebrates. As we’ve seen earlier, birds are highly modified diapsids, but rather than having two fenestrations or openings in their skulls behind the eye, the skulls of modern birds are so specialized that it is difficult to see any trace of the original diapsid condition.
Birds belong to a group of diapsids called the archosaurs, which includes three other groups: living crocodilians, pterosaurs, and dinosaurs. Overwhelming evidence shows that birds evolved within the clade Dinosauria, which is further subdivided into two groups, the Saurischia (“lizard hips”) and the Ornithischia (“bird hips”). Despite the names of these groups, it was not the bird-hipped dinosaurs that gave rise to modern birds. Rather, Saurischia diverged into two groups: One included the long-necked herbivorous dinosaurs, such as Apatosaurus. The second group, bipedal predators called theropods, gave rise to birds. This course of evolution is highlighted by numerous similarities between late (maniraptoran) theropod fossils and birds, specifically in the structure of the hip and wrist bones, as well as the presence of the wishbone, formed by the fusion of the clavicles.
The clade Neornithes includes the avian crown group, which comprises all living birds and the descendants from their most recent common maniraptoran ancestor. One well-known and important fossil of an animal that appears “intermediate” between dinosaurs and birds is Archaeopteryx ((Figure)), which is from the Jurassic period (200 to 145 MYA). Archaeopteryx has characteristics of both maniraptoran dinosaurs and modern birds. Some scientists propose classifying it as a bird, but others prefer to classify it as a dinosaur. Traits in skeletons of Archaeopteryx like those of a dinosaur included a jaw with teeth and a long bony tail. Like birds, it had feathers modified for flight, both on the forelimbs and on the tail, a trait associated only with birds among modern animals. Fossils of older feathered dinosaurs exist, but the feathers may not have had the characteristics of modern flight feathers.

The Evolution of Flight in Birds
There are two basic hypotheses that explain how flight may have evolved in birds: the arboreal (“tree”) hypothesis and the terrestrial (“land”) hypothesis. The arboreal hypothesis posits that tree-dwelling precursors to modern birds jumped from branch to branch using their feathers for gliding before becoming fully capable of flapping flight. In contrast to this, the terrestrial hypothesis holds that running (perhaps pursuing active prey such as small cursorial animals) was the stimulus for flight. In this scenario, wings could be used to capture prey and were preadapted for balance and flapping flight. Ostriches, which are large flightless birds, hold their wings out when they run, possibly for balance. However, this condition may represent a behavioral relict of the clade of flying birds that were their ancestors. It seems more likely that small feathered arboreal dinosaurs, were capable of gliding (and flapping) from tree to tree and branch to branch, improving the chances of escaping enemies, finding mates, and obtaining prey such as flying insects. This early flight behavior would have also greatly increased the opportunity for species dispersal.
Although we have a good understanding of how feathers and flight may have evolved, the question of how endothermy evolved in birds (and other lineages) remains unanswered. Feathers provide insulation, but this is only beneficial for thermoregulatory purposes if body heat is being produced internally. Similarly, internal heat production is only viable for the evolution of endothermy if insulation is present to retain that infrared energy. It has been suggested that one or the other—feathers or endothermy—evolved first in response to some other selective pressure (e.g., the ability to be active at night, provide camouflage, repel water, or serve as signals for mate selection). It seems probable that feathers and endothermy coevolved together, the improvement and evolutionary advancement of feathers reinforcing the evolutionary advancement of endothermy, and so on.
During the Cretaceous period (145 to 66 MYA), a group known as the Enantiornithes was the dominant bird type ((Figure)). Enantiornithes means “opposite birds,” which refers to the fact that certain bones of the shoulder are joined differently than the way the bones are joined in modern birds. Like Archaeopteryx, these birds retained teeth in their jaws, but did have a shortened tail, and at least some fossils have preserved “fans” of tail feathers. These birds formed an evolutionary lineage separate from that of modern birds, and they did not survive past the Cretaceous. Along with the Enantiornithes, however, another group of birds—the Ornithurae (“bird tails”), with a short, fused tail or pygostyle—emerged from the evolutionary line that includes modern birds. This clade was also present in the Cretaceous.
After the extinction of Enantiornithes, the Ornithurae became the dominant birds, with a large and rapid radiation occurring after the extinction of the dinosaurs during the Cenozoic era (66 MYA to the present). Molecular analysis based on very large data sets has produced our current understanding of the relationships among living birds. There are three major clades: the Paleognathae, the Galloanserae, and the Neoaves. The Paleognathae (“old jaw”) or ratites (polyphyletic) are a group of flightless birds including ostriches, emus, rheas, and kiwis. The Galloanserae include pheasants, ducks, geese and swans. The Neoaves (“new birds”) includes all other birds. The Neoaves themselves have been distributed among five clades:1 Strisores (nightjars, swifts, and hummingbirds), Columbaves (turacos, bustards, cuckoos, pigeons, and doves), Gruiformes (cranes), Aequorlitornithes (diving birds, wading birds, and shorebirds), and Inopinaves (a very large clade of land birds including hawks, owls, woodpeckers, parrots, falcons, crows, and songbirds). Despite the current classification scheme, it is important to understand that phylogenetic revisions, even for the extant birds, are still taking place.

Career Connection
VeterinarianVeterinarians are concerned with diseases, disorders, and injuries in animals, primarily vertebrates. They treat pets, livestock, and animals in zoos and laboratories. Veterinarians often treat dogs and cats, but also take care of birds, reptiles, rabbits, and other animals that are kept as pets. Veterinarians that work with farms and ranches care for pigs, goats, cows, sheep, and horses.
Veterinarians are required to complete a degree in veterinary medicine, which includes taking courses in comparative zoology, animal anatomy and physiology, microbiology, and pathology, among many other courses in chemistry, physics, and mathematics.
Veterinarians are also trained to perform surgery on many different vertebrate species, which requires an understanding of the vastly different anatomies of various species. For example, the stomach of ruminants like cows has four “compartments” versus one compartment for non-ruminants. As we have seen, birds also have unique anatomical adaptations that allow for flight, which requires additional training and care.
Some veterinarians conduct research in academic settings, broadening our knowledge of animals and medical science. One area of research involves understanding the transmission of animal diseases to humans, called zoonotic diseases. For example, one area of great concern is the transmission of the avian flu virus to humans. One type of avian flu virus, H5N1, is a highly pathogenic strain that has been spreading in birds in Asia, Europe, Africa, and the Middle East. Although the virus does not cross over easily to humans, there have been cases of bird-to-human transmission. More research is needed to understand how this virus can cross the species barrier and how its spread can be prevented.
Section Summary
Birds are the most speciose group of land vertebrates and display a number of adaptations related to their ability to fly, which were first present in their therapod (maniraptoran) ancestors. Birds are endothermic (and homeothermic), meaning they have a very high metabolism that produces a considerable amount of heat, as well as structures such as feathers that allow them to retain their own body heat. These adaptations are used to regulate their internal temperature, making it largely independent of ambient thermal conditions.
Birds have feathers, which allow for insulation and flight, as well as for mating and warning signals. Flight feathers have a broad and continuously curved vane that produces lift. Some birds have pneumatic bones containing air spaces that are sometimes connected to air sacs in the body cavity. Airflow through bird lungs travels in one direction, creating a counter-current gas exchange with the blood.
Birds are highly modified diapsids and belong to a group called the archosaurs. Within the archosaurs, birds are most likely evolved from theropod (maniraptoran) dinosaurs. One of the oldest known fossils (and best known) of a “dinosaur-bird” is that of Archaeopteryx, which is dated from the Jurassic period. Modern birds are now classified into three groups: Paleognathae, Galloanserae, and Neoaves.
Footnotes
- 1 Prum, RO et al. 2015. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526: 569 – 573. http://dx.doi.org/10.1038/nature15697
Glossary
- Archaeopteryx
- transition species from dinosaur to bird from the Jurassic period
- contour feather
- feather that creates an aerodynamic surface for efficient flight
- down feather
- feather specialized for insulation
- Enantiornithes
- dominant bird group during the Cretaceous period
- flight feather
- feather specialized for flight
- furcula
- wishbone formed by the fusing of the clavicles
- Neognathae
- birds other than the Paleognathae
- Neornithes
- modern birds
- Paleognathae
- ratites; flightless birds, including ostriches and emus
- pneumatic bone
- air-filled bone
- primary feather
- feather located at the tip of the wing that provides thrust
- secondary feather
- feather located at the base of the wing that provides lift
- theropod
- dinosaur group ancestral to birds
