
Animal Diversity
12.1 – Features of the Animal Kingdom
Learning Objectives
By the end of this section, you will be able to do the following:
- List the features that distinguish the kingdom Animalia from other kingdoms
- Explain the processes of animal reproduction and embryonic development
- Describe the roles that Hox genes play in development
Two different groups within the Domain Eukaryota have produced complex multicellular organisms: The plants arose within the Archaeplastida, whereas the animals (and their close relatives, the fungi) arose within the Opisthokonta. However, plants and animals not only have different life styles, they also have different cellular histories as eukaryotes. The opisthokonts share the possession of a single posterior flagellum in flagellated cells, e.g., sperm cells.
Most animals also share other features that distinguish them from organisms in other kingdoms. All animals require a source of food and are therefore heterotrophic, ingesting other living or dead organisms. This feature distinguishes them from autotrophic organisms, such as most plants, which synthesize their own nutrients through photosynthesis. As heterotrophs, animals may be carnivores, herbivores, omnivores, or parasites ((Figure)a,b). As with plants, almost all animals have a complex tissue structure with differentiated and specialized tissues. The necessity to collect food has made most animals motile, at least during certain life stages. The typical life cycle in animals is diplontic (like you, the diploid state is multicellular, whereas the haploid state is gametic, such as sperm or egg). We should note that the alternation of generations characteristic of the land plants is typically not found in animals. In animals whose life histories include several to multiple body forms (e.g., insect larvae or the medusae of some Cnidarians), all body forms are diploid. Animal embryos pass through a series of developmental stages that establish a determined and fixed body plan. The body plan refers to the morphology of an animal, determined by developmental cues.

Complex Tissue Structure
Many of the specialized tissues of animals are associated with the requirements and hazards of seeking and processing food. This explains why animals typically have evolved special structures associated with specific methods of food capture and complex digestive systems supported by accessory organs. Sensory structures help animals navigate their environment, detect food sources (and avoid becoming a food source for other animals!). Movement is driven by muscle tissue attached to supportive structures like bone or chitin, and is coordinated by neural communication. Animal cells may also have unique structures for intercellular communication (such as gap junctions). The evolution of nerve tissues and muscle tissues has resulted in animals’ unique ability to rapidly sense and respond to changes in their environment. This allows animals to survive in environments where they must compete with other species to meet their nutritional demands.
The tissues of animals differ from those of the other major multicellular eukaryotes, plants and fungi, because their cells don’t have cell walls. However, cells of animal tissues may be embedded in an extracellular matrix (e.g., mature bone cells reside within a mineralized organic matrix secreted by the cells). In vertebrates, bone tissue is a type of connective tissue that supports the entire body structure. The complex bodies and activities of vertebrates demand such supportive tissues. Epithelial tissues cover and protect both external and internal body surfaces, and may also have secretory functions. Epithelial tissues include the epidermis of the integument, the lining of the digestive tract and trachea, as well as the layers of cells that make up the ducts of the liver and glands of advanced animals, for example. The different types of tissues in true animals are responsible for carrying out specific functions for the organism. This differentiation and specialization of tissues is part of what allows for such incredible animal diversity.
Just as there are multiple ways to be a eukaryote, there are multiple ways to be a multicellular animal. The animal kingdom is currently divided into five monophyletic clades: Parazoa or Porifera (sponges), Placozoa (tiny parasitic creatures that resemble multicellular amoebae), Cnidaria (jellyfish and their relatives), Ctenophora (the comb jellies), and Bilateria (all other animals). The Placozoa (“flat animal”) and Parazoa (“beside animal”) do not have specialized tissues derived from germ layers of the embryo; although they do possess specialized cells that act functionally like tissues. The Placozoa have only four cell types, while the sponges have nearly two dozen. The three other clades do include animals with specialized tissues derived from the germ layers of the embryo. In spite of their superficial similarity to Cnidarian medusae, recent molecular studies indicate that the Ctenophores are only distantly related to the Cnidarians, which together with the Bilateria constitute the Eumetazoa (“true animals”). When we think of animals, we usually think of Eumetazoa, since most animals fall into this category.
Link to Learning
Watch a presentation by biologist E.O. Wilson on the importance of diversity.
Animal Reproduction and Development
Most animals are diploid organisms, meaning that their body (somatic) cells are diploid and haploid reproductive (gamete) cells are produced through meiosis. Some exceptions exist: for example, in bees, wasps, and ants, the male is haploid because it develops from unfertilized eggs. Most animals undergo sexual reproduction. However, a few groups, such as cnidarians, flatworms, and roundworms, may also undergo asexual reproduction, in which offspring originate from part of the parental body.
Processes of Animal Reproduction and Embryonic Development
During sexual reproduction, the haploid gametes of the male and female individuals of a species combine in a process called fertilization. Typically, both male and female gametes are required: the small, motile male sperm fertilizes the typically much larger, sessile female egg. This process produces a diploid fertilized egg called a zygote.
Some animal species—including sea stars and sea anemones—are capable of asexual reproduction. The most common forms of asexual reproduction for stationary aquatic animals include budding and fragmentation, where part of a parent individual can separate and grow into a new individual. This type of asexual reproduction produces genetically identical offspring, which would appear to be disadvantageous from the perspective of evolutionary adaptability, simply because of the potential buildup of deleterious mutations.
In contrast, a form of uniparental reproduction found in some insects and a few vertebrates is called parthenogenesis (or “virgin beginning”). In this case, progeny develop from a gamete, but without fertilization. Because of the nutrients stored in eggs, only females produce parthenogenetic offspring. In some insects, unfertilized eggs develop into new male offspring. This type of sex determination is called haplodiploidy, since females are diploid (with both maternal and paternal chromosomes) and males are haploid (with only maternal chromosomes). A few vertebrates, e.g., some fish, turkeys, rattlesnakes, and whiptail lizards, are also capable of parthenogenesis. In the case of turkeys and rattlesnakes, parthenogenetically reproducing females also produce only male offspring, but not because the males are haploid. In birds and rattlesnakes, the female is the heterogametic (ZW) sex, so the only surviving progeny of post-meiotic parthenogenesis would be ZZ males. In the whiptail lizards, on the other hand, only female progeny are produced by parthenogenesis. These animals may not be identical to their parent, although they have only maternal chromosomes. However, for animals that are limited in their access to mates, uniparental reproduction can ensure genetic propagation.
In animals, the zygote progresses through a series of developmental stages, during which primary germ layers (ectoderm, endoderm, and mesoderm) are established and reorganize to form an embryo. During this process, animal tissues begin to specialize and organize into organs and organ systems, determining their future morphology and physiology.
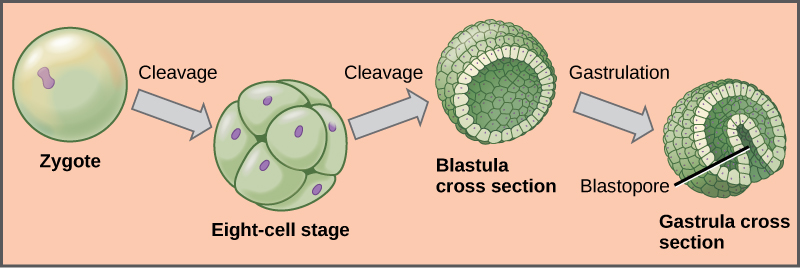
Animal development begins with cleavage, a series of mitotic cell divisions, of the zygote ((Figure)). Cleavage differs from somatic cell division in that the egg is subdivided by successive cleavages into smaller and smaller cells, with no actual cell growth. The cells resulting from subdivision of the material of the egg in this way are called blastomeres. Three cell divisions transform the single-celled zygote into an eight-celled structure. After further cell division and rearrangement of existing cells, a solid morula is formed, followed by a hollow structure called a blastula. The blastula is hollow only in invertebrates whose eggs have relatively small amounts of yolk. In very yolky eggs of vertebrates, the yolk remains undivided, with most cells forming an embryonic layer on the surface of the yolk (imagine a chicken embryo growing over the egg’s yolk), which serve as food for the developing embryo.
Further cell division and cellular rearrangement leads to a process called gastrulation. Gastrulation results in two important events: the formation of the primitive gut (archenteron) or digestive cavity, and the formation of the embryonic germ layers, as we have discussed above. These germ layers are programmed to develop into certain tissue types, organs, and organ systems during a process called organogenesis. Diploblastic organisms have two germ layers, endoderm and ectoderm. Endoderm forms the wall of the digestive tract, and ectoderm covers the surface of the animal. In triploblastic animals, a third layer forms: mesoderm, which differentiates into various structures between the ectoderm and endoderm, including the lining of the body cavity.
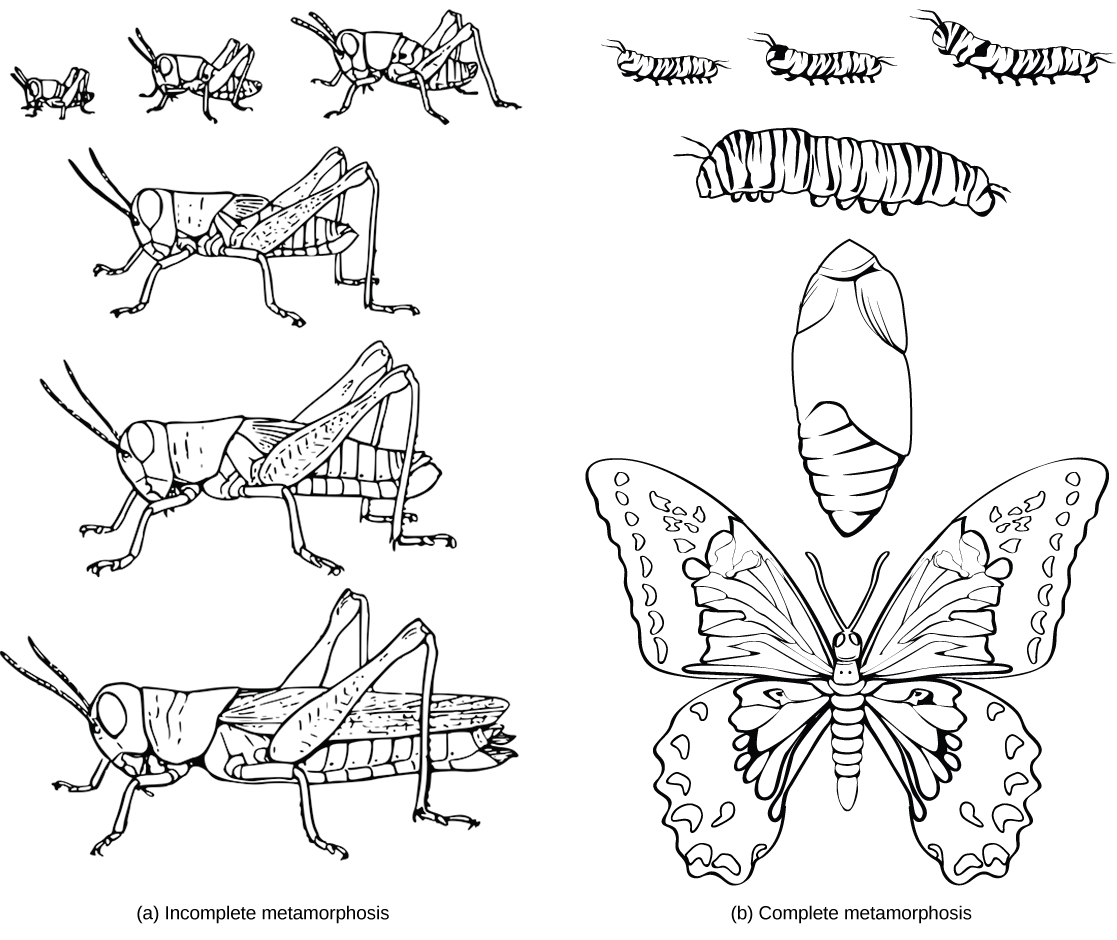
Some animals produce larval forms that are different from the adult. In insects with incomplete metamorphosis, such as grasshoppers, the young resemble wingless adults, but gradually produce larger and larger wing buds during successive molts, until finally producing functional wings and sex organs during the last molt. Other animals, such as some insects and echinoderms, undergo complete metamorphosis in which the embryo develops into one or more feeding larval stages that may differ greatly in structure and function from the adult ((Figure)). The adult body then develops from one or more regions of larval tissue. For animals with complete metamorphosis, the larva and the adult may have different diets, limiting competition for food between them. Regardless of whether a species undergoes complete or incomplete metamorphosis, the series of developmental stages of the embryo remains largely the same for most members of the animal kingdom.
Link to Learning
Watch the following video to see how human embryonic development (after the blastula and gastrula stages of development) reflects evolution.
The Role of Homeobox (Hox) Genes in Animal Development
Since the early nineteenth century, scientists have observed that many animals, from the very simple to the complex, shared similar embryonic morphology and development. Surprisingly, a human embryo and a frog embryo, at a certain stage of embryonic development, look remarkably alike! For a long time, scientists did not understand why so many animal species looked similar during embryonic development but were very different as adults. They wondered what dictated the developmental direction that a fly, mouse, frog, or human embryo would take. Near the end of the twentieth century, a particular class of genes was discovered that had this very job. These genes that determine animal structure are called “homeotic genes,” and they contain DNA sequences called homeoboxes. Genes with homeoboxes encode protein transcription factors. One group of animal genes containing homeobox sequences is specifically referred to as Hox genes. This cluster of genes is responsible for determining the general body plan, such as the number of body segments of an animal, the number and placement of appendages, and animal head-tail directionality. The first Hox genes to be sequenced were those from the fruit fly (Drosophila melanogaster). A single Hox mutation in the fruit fly can result in an extra pair of wings or even legs growing from the head in place of antennae (this is because antennae and legs are embryologic homologous structures and their appearance as antennae or legs is dictated by their origination within specific body segments of the head and thorax during development). Now, Hox genes are known from virtually all other animals as well.
While there are a great many genes that play roles in the morphological development of an animal, including other homeobox-containing genes, what makes Hox genes so powerful is that they serve as “master control genes” that can turn on or off large numbers of other genes. Hox genes do this by encoding transcription factors that control the expression of numerous other genes. Hox genes are homologous across the animal kingdom, that is, the genetic sequences of Hox genes and their positions on chromosomes are remarkably similar across most animals because of their presence in a common ancestor, from worms to flies, mice, and humans ((Figure)). In addition, the order of the genes reflects the anterior-posterior axis of the animal’s body. One of the contributions to increased animal body complexity is that Hox genes have undergone at least two and perhaps as many as four duplication events during animal evolution, with the additional genes allowing for more complex body types to evolve. All vertebrates have four (or more) sets of Hox genes, while invertebrates have only one set.
Visual Connection
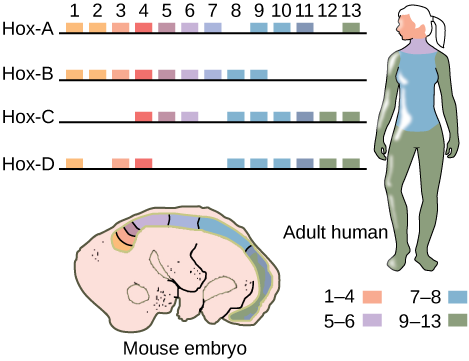
If a Hox 13 gene in a mouse was replaced with a Hox 1 gene, how might this alter animal development?
Two of the five clades within the animal kingdom do not have Hox genes: the Ctenophora and the Porifera. In spite of the superficial similarities between the Cnidaria and the Ctenophora, the Cnidaria have a number of Hox genes, but the Ctenophora have none. The absence of Hox genes from the ctenophores has led to the suggestion that they might be “basal” animals, in spite of their tissue differentiation. Ironically, the Placozoa, which have only a few cell types, do have at least one Hox gene. The presence of a Hox gene in the Placozoa, in addition to similarities in the genomic organization of the Placozoa, Cnidaria and Bilateria, has led to the inclusion of the three groups in a “Parahoxozoa” clade. However, we should note that at this time the reclassification of the Animal Kingdom is still tentative and requires much more study.
Section Summary
Animals constitute an incredibly diverse kingdom of organisms. Although animals range in complexity from simple sea sponges to human beings, most members of the animal kingdom share certain features. Animals are eukaryotic, multicellular, heterotrophic organisms that ingest their food and usually develop into motile creatures with a fixed body plan. A major characteristic unique to the animal kingdom is the presence of differentiated tissues, such as nerve, muscle, and connective tissues, which are specialized to perform specific functions. Most animals undergo sexual reproduction, leading to a series of developmental embryonic stages that are relatively similar across the animal kingdom. A class of transcriptional control genes called Hox genes directs the organization of the major animal body plans, and these genes are strongly homologous across the animal kingdom.
Glossary
- blastula
- 16–32 cell stage of development of an animal embryo
- body plan
- morphology or defining shape of an organism
- cleavage
- cell divisions subdividing a fertilized egg (zygote) to form a multicellular embryo
- gastrula
- stage of animal development characterized by the formation of the digestive cavity
- germ layer
- collection of cells formed during embryogenesis that will give rise to future body tissues, more pronounced in vertebrate embryogenesis
- Hox gene
- (also, homeobox gene) master control gene that can turn on or off large numbers of other genes during embryogenesis
- organogenesis
- formation of organs in animal embryogenesis
