
Animal Diversity
13.2 – Phylum Cnidaria
Learning Objectives
By the end of this section, you will be able to do the following:
- Compare structural and organization characteristics of Porifera and Cnidaria
- Describe the progressive development of tissues and their relevance to animal complexity
- Identify the two general body forms found in the Cnidaria
- Describe the identifying features of the major cnidarian classes
Phylum Cnidaria includes animals that exhibit radial or biradial symmetry and are diploblastic, meaning that they develop from two embryonic layers, ectoderm and endoderm. Nearly all (about 99 percent) cnidarians are marine species.
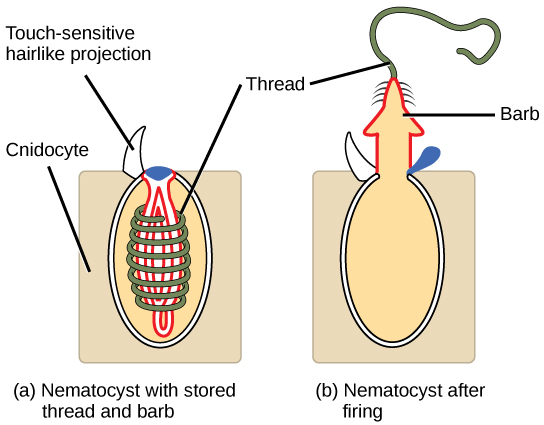
Whereas the defining cell type for the sponges is the choanocyte, the defining cell type for the cnidarians is the cnidocyte, or stinging cell. These cells are located around the mouth and on the tentacles, and serve to capture prey or repel predators. Cnidocytes have large stinging organelles called nematocysts, which usually contain barbs at the base of a long coiled thread. The outer wall of the cell has a hairlike projection called a cnidocil, which is sensitive to tactile stimulation. If the cnidocils are touched, the hollow threads evert with enormous acceleration, approaching 40,000 times that of gravity. The microscopic threads then either entangle the prey or instantly penetrate the flesh of the prey or predator, releasing toxins (including neurotoxins and pore-forming toxins that can lead to cell lysis) into the target, thereby immobilizing it or paralyzing it (see (Figure)).
Link to Learning
View this video animation showing two anemones engaged in a battle.
Two distinct body plans are found in Cnidarians: the polyp or tuliplike “stalk” form and the medusa or “bell” form. ((Figure)). An example of the polyp form is found in the genus Hydra, whereas the most typical form of medusa is found in the group called the “sea jellies” (jellyfish). Polyp forms are sessile as adults, with a single opening (the mouth/anus) to the digestive cavity facing up with tentacles surrounding it. Medusa forms are motile, with the mouth and tentacles hanging down from an umbrella-shaped bell.
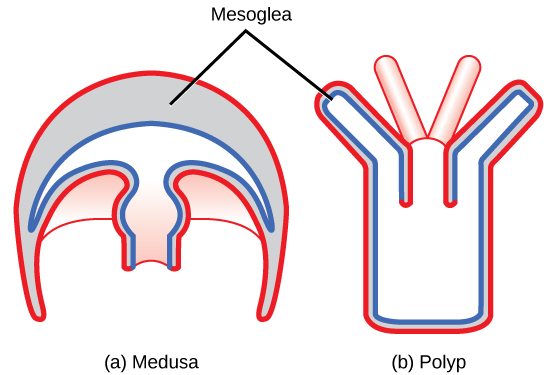
Some cnidarians are dimorphic, that is, they exhibit both body plans during their life cycle. In these species, the polyp serves as the asexual phase, while the medusa serves as the sexual stage and produces gametes. However, both body forms are diploid.
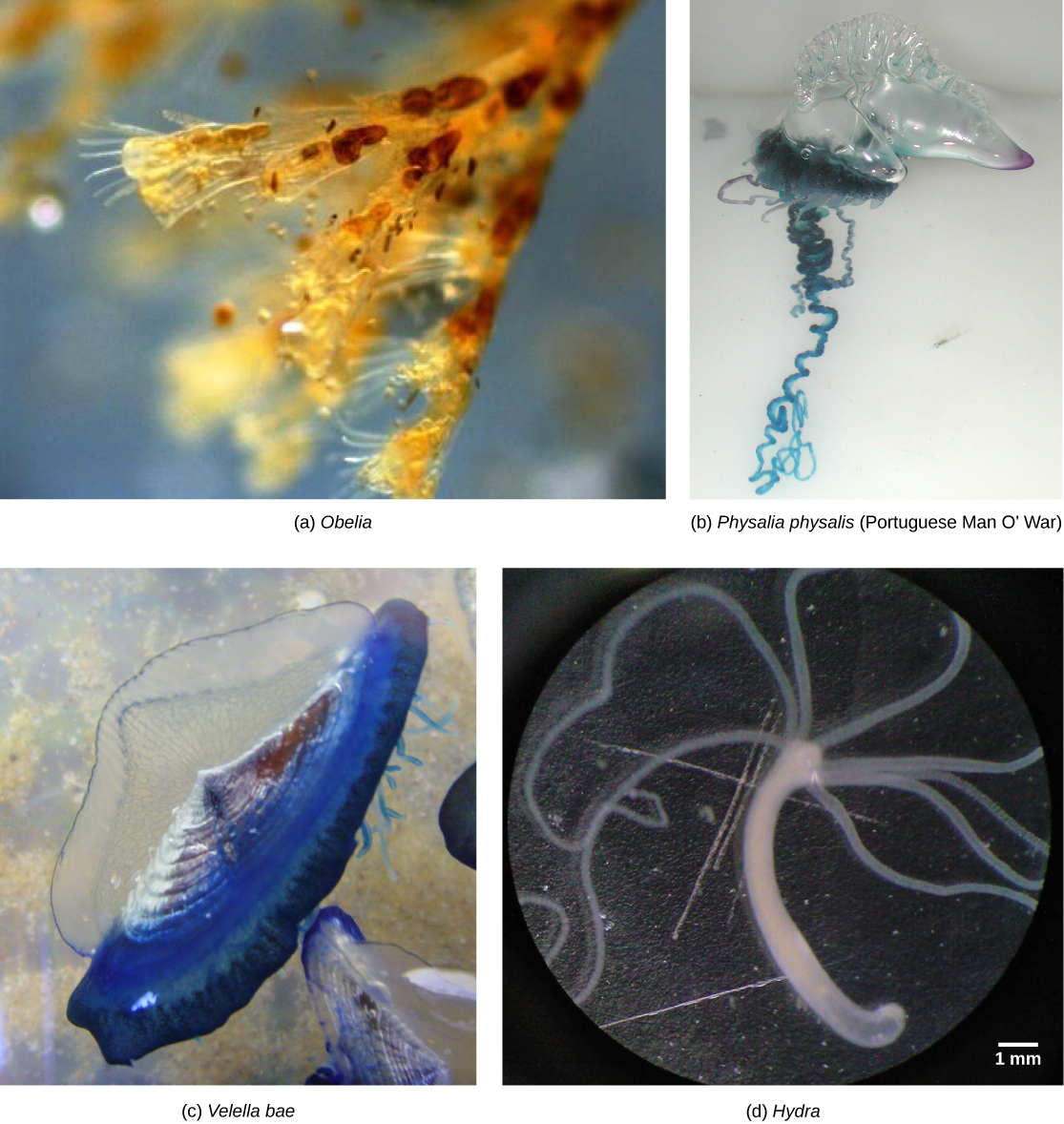
An example of cnidarian dimorphism can be seen in the colonial hydroid Obelia. The sessile asexual colony has two types of polyps, shown in (Figure). The first is the gastrozooid, which is adapted for capturing prey and feeding. In Obelia, all polyps are connected through a common digestive cavity called a coenosarc. The other type of polyp is the gonozooid, adapted for the asexual budding and the production of sexual medusae. The reproductive buds from the gonozooid break off and mature into free-swimming medusae, which are either male or female (dioecious). Each medusa has either several testes or several ovaries in which meiosis occurs to produce sperm or egg cells. Interestingly, the gamete-producing cells do not arise within the gonad itself, but migrate into it from the tissues in the gonozooid. This separate origin of gonad and gametes is common throughout the eumetazoa. The gametes are released into the surrounding water, and after fertilization, the zygote develops into a blastula, which soon develops into a ciliated, bilaterally symmetrical planula larva. The planula swims freely for a while, but eventually attaches to a substrate and becomes a single polyp, from which a new colony of polyps is formed by budding.

Link to Learning
Click here to follow an Obelia life cycle animation and quiz.
All cnidarians are diploblastic and thus have two “epithelial” layers in the body that are derived from the endoderm and ectoderm of the embryo. The outer layer (from ectoderm) is called the epidermis and lines the outside of the animal, whereas the inner layer (from endoderm) is called the gastrodermis and lines the digestive cavity. In the planula larva, a layer of ectoderm surrounds a solid mass of endoderm, but as the polyp develops, the digestive or gastrovascular cavity opens within the endoderm. A non-living, jelly-like mesoglea lies between these two epithelial layers. In terms of cellular complexity, cnidarians show the presence of differentiated cell types in each tissue layer, such as nerve cells, contractile epithelial cells, enzyme-secreting cells, and nutrient-absorbing cells, as well as the presence of intercellular connections. However, with a few notable exceptions such as statocysts and rhopalia (see below), the development of organs or organ systems is not advanced in this phylum.
The nervous system is rudimentary, with nerve cells organized in a network scattered across the body. This nerve net may show the presence of groups of cells that form nerve plexi (singular: plexus) or nerve cords. Organization of the nervous system in the motile medusa is more complex than that of the sessile polyp, with a nerve ring around the edge of the medusa bell that controls the action of the tentacles. Cnidarian nerve cells show mixed characteristics of motor and sensory neurons. The predominant signaling molecules in these primitive nervous systems are peptides, which perform both excitatory and inhibitory functions. Despite the simplicity of the nervous system, it is remarkable that it coordinates the complicated movement of the tentacles, the drawing of captured prey to the mouth, the digestion of food, and the expulsion of waste.
The gastrovascular cavity has only one opening that serves as both a mouth and an anus; this arrangement is called an incomplete digestive system. In the gastrovascular cavity, extracellular digestion occurs as food is taken into the gastrovascular cavity, enzymes are secreted into the cavity, and the cells lining the cavity absorb nutrients. However, some intracellular digestion also occurs. The gastrovascular cavity distributes nutrients throughout the body of the animal, with nutrients passing from the digestive cavity across the mesoglea to the epidermal cells. Thus, this cavity serves both digestive and circulatory functions.
Cnidarian cells exchange oxygen and carbon dioxide by diffusion between cells in the epidermis and water in the environment, and between cells in the gastrodermis and water in the gastrovascular cavity. The lack of a circulatory system to move dissolved gases limits the thickness of the body wall and necessitates a non-living mesoglea between the layers. In the cnidarians with a thicker mesoglea, a number of canals help to distribute both nutrients and gases. There is neither an excretory system nor organs, and nitrogenous wastes simply diffuse from the cells into the water outside the animal or into the gastrovascular cavity.
The phylum Cnidaria contains about 10,000 described species divided into two monophyletic clades: the Anthozoa and the Medusozoa. The Anthozoa include the corals, sea fans, sea whips, and the sea anemones. The Medusozoa include several classes of Cnidaria in two clades: The Hydrozoa include sessile forms, some medusoid forms, and swimming colonial forms like the Portuguese man-of-war. The other clade contains various types of jellies including both Scyphozoa and Cubozoa. The Anthozoa contain only sessile polyp forms, while the Medusozoa include species with both polyp and medusa forms in their life cycle.
Class Anthozoa
The class Anthozoa (“flower animals”) includes sea anemones ((Figure)), sea pens, and corals, with an estimated number of 6,100 described species. Sea anemones are usually brightly colored and can attain a size of 1.8 to 10 cm in diameter. Individual animals are cylindrical in shape and are attached directly to a substrate.
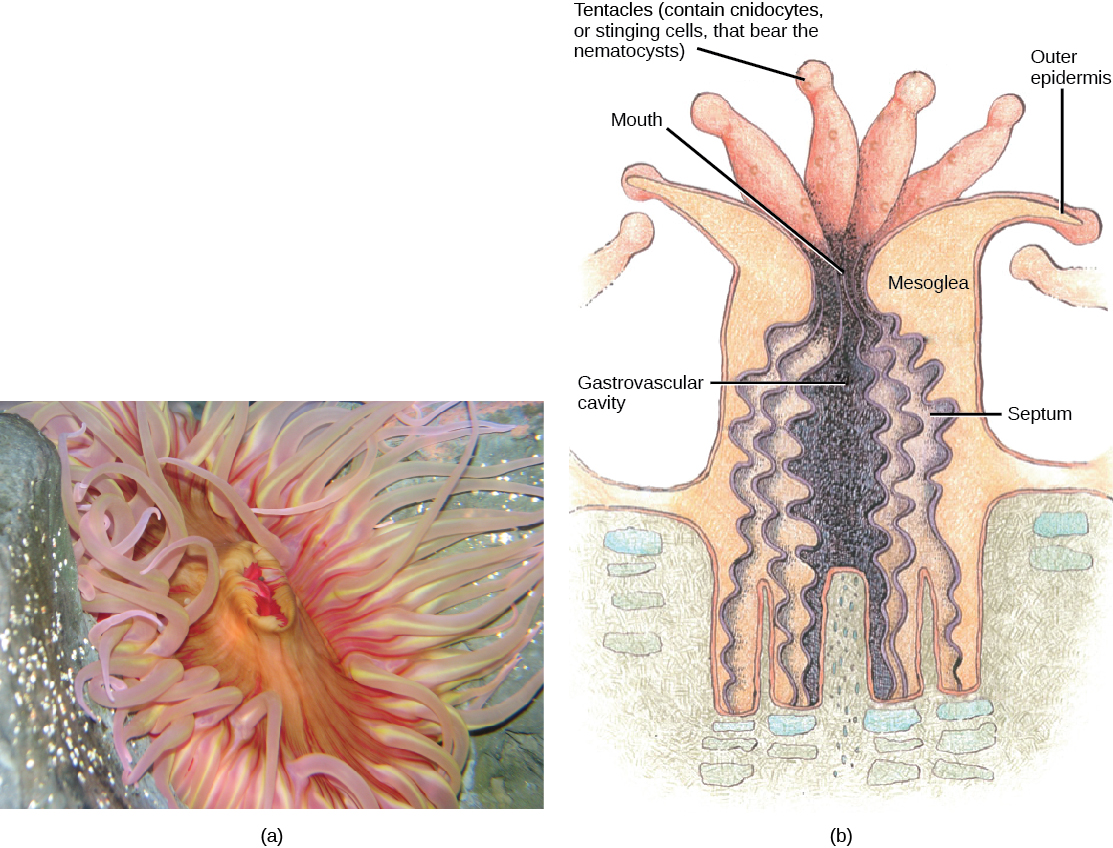
The mouth of a sea anemone is surrounded by tentacles that bear cnidocytes. The slit-like mouth opening and flattened pharynx are lined with ectoderm. This structure of the pharynx makes anemones bilaterally symmetrical. A ciliated groove called a siphonoglyph is found on two opposite sides of the pharynx and directs water into it. The pharynx is the muscular part of the digestive system that serves to ingest as well as egest food, and may extend for up to two-thirds the length of the body before opening into the gastrovascular cavity. This cavity is divided into several chambers by longitudinal septa called mesenteries. Each mesentery consists of a fold of gastrodermal tissue with a layer of mesoglea between the sheets of gastrodermis. Mesenteries do not divide the gastrovascular cavity completely, and the smaller cavities coalesce at the pharyngeal opening. The adaptive benefit of the mesenteries appears to be an increase in surface area for absorption of nutrients and gas exchange, as well as additional mechanical support for the body of the anemone.
Sea anemones feed on small fish and shrimp, usually by immobilizing their prey with nematocysts. Some sea anemones establish a mutualistic relationship with hermit crabs when the crab seizes and attaches them to their shell. In this relationship, the anemone gets food particles from prey caught by the crab, and the crab is protected from the predators by the stinging cells of the anemone. Some species of anemone fish, or clownfish, are also able to live with sea anemones because they build up an acquired immunity to the toxins contained within the nematocysts and also secrete a protective mucus that prevents them from being stung.
The structure of coral polyps is similar to that of anemones, although the individual polyps are usually smaller and part of a colony, some of which are massive and the size of small buildings. Coral polyps feed on smaller planktonic organisms, including algae, bacteria, and invertebrate larvae. Some anthozoans have symbiotic associations with dinoflagellate algae called zooxanthellae. The mutually beneficial relationship between zooxanthellae and modern corals—which provides the algae with shelter—gives coral reefs their colors and supplies both organisms with nutrients. This complex mutualistic association began more than 210 million years ago, according to a new study by an international team of scientists. That this symbiotic relationship arose during a time of massive worldwide coral-reef expansion suggests that the interconnection of algae and coral is crucial for the health of coral reefs, which provide habitat for roughly one-fourth of all marine life. Reefs are threatened by a trend in ocean warming that has caused corals to expel their zooxanthellae algae and turn white, a process called coral bleaching.
Anthozoans remain polypoid (note that this term is easily confused with “polyploid”) throughout their lives and can reproduce asexually by budding or fragmentation, or sexually by producing gametes. Male or female gametes produced by a polyp fuse to give rise to a free-swimming planula larva. The larva settles on a suitable substratum and develops into a sessile polyp.
Class Scyphozoa
Class Scyphozoa (“cup animals”) includes all (and only) the marine jellies, with about 200 known species. The medusa is the prominent stage in the life cycle, although there is a polyp stage in the life cycle of most species. Most jellies range from 2 to 40 cm in length but the largest scyphozoan species, Cyanea capillata, can reach a size of two meters in diameter. Scyphozoans display a characteristic bell-like morphology ((Figure)).
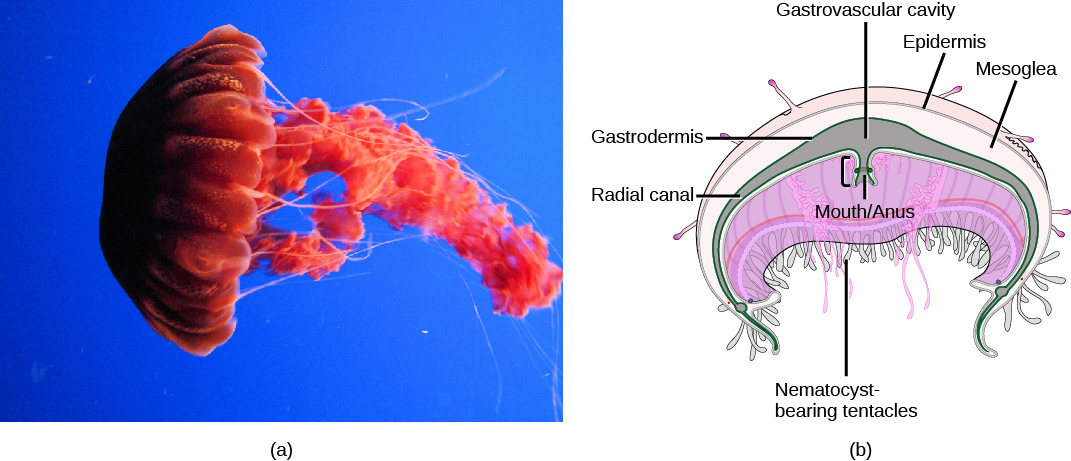
In the sea jelly, a mouth opening is present on the underside of the animal, surrounded by hollow tentacles bearing nematocysts. Scyphozoans live most of their life cycle as free-swimming, solitary carnivores. The mouth leads to the gastrovascular cavity, which may be sectioned into four interconnected sacs, called diverticuli. In some species, the digestive system may branch further into radial canals. Like the septa in anthozoans, the branched gastrovascular cells serve two functions: to increase the surface area for nutrient absorption and diffusion, and to support the body of the animal.
In scyphozoans, nerve cells are organized in a nerve net that extends over the entire body, with a nerve ring around the edge of the bell. Clusters of sensory organs called rhopalia may be present in pockets in the edge of the bell. Jellies have a ring of muscles lining the dome of the body, which provides the contractile force required to swim through water, as well as to draw in food from the water as they swim. Scyphozoans have separate sexes. The gonads are formed from the gastrodermis and gametes are expelled through the mouth. Planula larvae are formed by external fertilization; they settle on a substratum in a polypoid form. These polyps may bud to form additional polyps or begin immediately to produce medusa buds. In a few species, the planula larva may develop directly into the medusa. The life cycle ((Figure)) of most scyphozoans includes both sexual medusoid and asexual polypoid body forms.
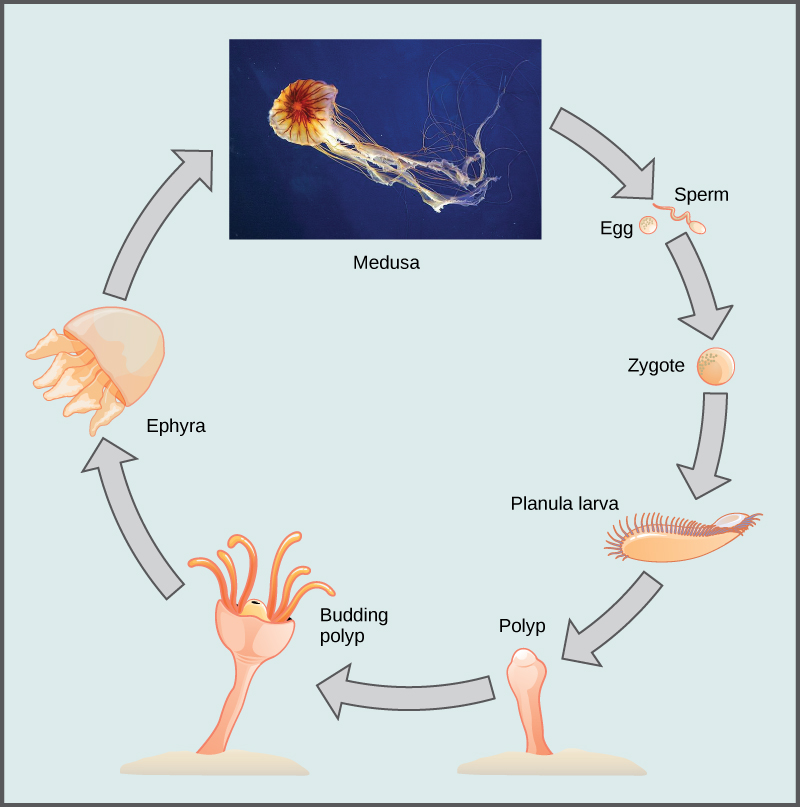
Class Cubozoa
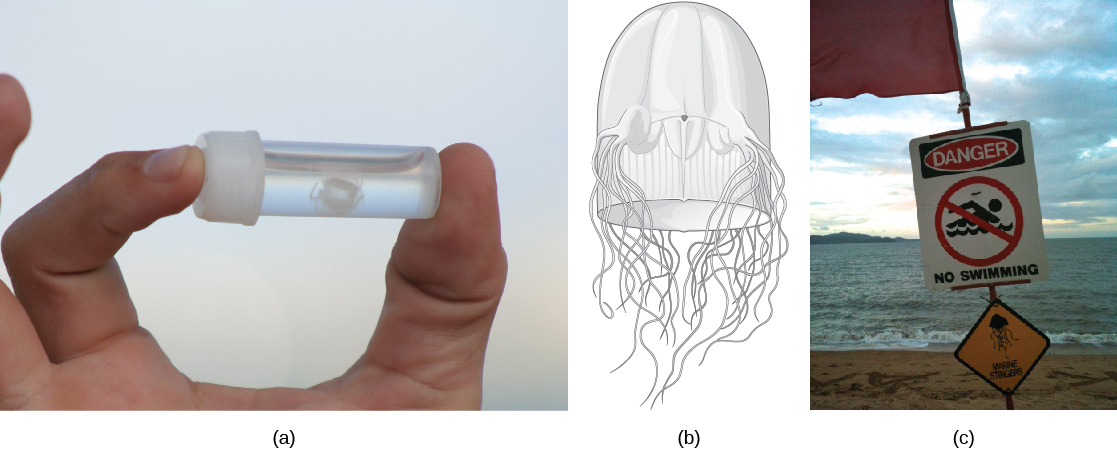
This class includes jellies that have a box-shaped medusa, or a bell that is square in cross-section, and are colloquially known as “box jellyfish.” These species may achieve sizes of 15 to 25 cm, but typically members of the Cubozoa are not as large as those of the Scyphozoa. However, cubozoans display overall morphological and anatomical characteristics that are similar to those of the scyphozoans. A prominent difference between the two classes is the arrangement of tentacles. The cubozoans contain muscular pads called pedalia at the corners of the square bell canopy, with one or more tentacles attached to each pedalium. In some cases, the digestive system may extend into the pedalia. Nematocysts may be arranged in a spiral configuration along the tentacles; this arrangement helps to effectively subdue and capture prey. Cubozoans include the most venomous of all the cnidarians ((Figure)).
These animals are unusual in having image-forming eyes, including a cornea, lens, and retina. Because these structures are made from a number of interactive tissues, they can be called true organs. Eyes are located in four clusters between each pair of pedalia. Each cluster consists of four simple eye spots plus two image-forming eyes oriented in different directions. How images formed by these very complex eyes are processed remains a mystery, since cubozoans have extensive nerve nets but no distinct brain. Nontheless, the presence of eyes helps the cubozoans to be active and effective hunters of small marine animals like worms, arthropods, and fish.
Cubozoans have separate sexes and fertilization occurs inside the female. Planula larvae may develop inside the female or be released, depending on species. Each planula develops into a polyp. These polyps may bud to form more polyps to create a colony; each polyp then transforms into a single medusa.
Class Hydrozoa
Hydrozoa is a diverse group that includes nearly 3,200 species; most are marine, although some freshwater species are known ((Figure)). Most species exhibit both polypoid and medusoid forms in their lifecycles, although the familiar Hydra has only the polyp form. The medusoid form has a muscular veil or velum below the margin of the bell and for this reason is called a hydromedusa. In contrast, the medusoid form of Scyphozoa lacks a velum and is termed a scyphomedusa.
The polyp form in these animals often shows a cylindrical morphology with a central gastrovascular cavity lined by the gastrodermis. The gastrodermis and epidermis have a simple layer of mesoglea sandwiched between them. A mouth opening, surrounded by tentacles, is present at the oral end of the animal. Many hydrozoans form sessile, branched colonies of specialized polyps that share a common, branching gastrovascular cavity (coenosarc), such as is found in the colonial hydroid Obelia.
Free-floating colonial species called siphonophores contain both medusoid and polypoid individuals that are specialized for feeding, defense, or reproduction. The distinctive rainbow-hued float of the Portuguese man o’ war (Physalia physalis) creates a pneumatophore with which it regulates buoyancy by filling and expelling carbon monoxide gas. At first glance, these complex superorganisms appear to be a single organism; but the reality is that even the tentacles are actually composed of zooids laden with nematocysts. Thus, although it superficially resembles a typical medusozoan jellyfish, P. physalis is a free-floating hydrozoan colony; each specimen is made up of many hundreds of organisms, each specialized for a certain function, including motility and buoyancy, feeding, reproduction and defense. Although they are carnivorous and feed on many soft bodied marine animals, P. physalis lack stomachs and instead have specialized polyps called gastrozooids that they use to digest their prey in the open water.
Physalia has male and female colonies, which release their gametes into the water. The zygote develops into a single individual, which then buds asexually to form a new colony. Siphonophores include the largest known floating cnidarian colonies such as Praya dubia, whose chain of zoids can get up to 50 meters (165 feet) long. Other hydrozoan species are solitary polyps (Hydra) or solitary hydromedusae (Gonionemus). One defining characteristic shared by the hydrozoans is that their gonads are derived from epidermal tissue, whereas in all other cnidarians they are derived from gastrodermal tissue.
Section Summary
Cnidarians represent a more complex level of organization than Porifera. They possess outer and inner tissue layers that sandwich a noncellular mesoglea between them. Cnidarians possess a well-formed digestive system and carry out extracellular digestion in a digestive cavity that extends through much of the animal. The mouth is surrounded by tentacles that contain large numbers of cnidocytes—specialized cells bearing nematocysts used for stinging and capturing prey as well as discouraging predators. Cnidarians have separate sexes and many have a lifecycle that involves two distinct morphological forms—medusoid and polypoid—at various stages in their life cycles. In species with both forms, the medusa is the sexual, gamete-producing stage and the polyp is the asexual stage. Cnidarian species include individual or colonial polypoid forms, floating colonies, or large individual medusa forms (sea jellies).
Glossary
- Cnidaria
- phylum of animals that are diploblastic and have radial symmetry
- cnidocyte
- specialized stinging cell found in Cnidaria
- epidermis
- outer layer (from ectoderm) that lines the outside of the animal
- extracellular digestion
- food is taken into the gastrovascular cavity, enzymes are secreted into the cavity, and the cells lining the cavity absorb nutrients
- gastrodermis
- inner layer (from endoderm) that lines the digestive cavity
- gastrovascular cavity
- opening that serves as both a mouth and an anus, which is termed an incomplete digestive system
- medusa
- free-floating cnidarian body plan with mouth on underside and tentacles hanging down from a bell
- mesoglea
- non-living, gel-like matrix present between ectoderm and endoderm in cnidarians
- nematocyst
- harpoon-like organelle within cnidocyte with pointed projectile and poison to stun and entangle prey
- polyp
- stalk-like sessile life form of a cnidarians with mouth and tentacles facing upward, usually sessile but may be able to glide along surface
- polymorphic
- possessing multiple body plans within the lifecycle of a group of organisms
- siphonophore
- tubular structure that serves as an inlet for water into the mantle cavity
